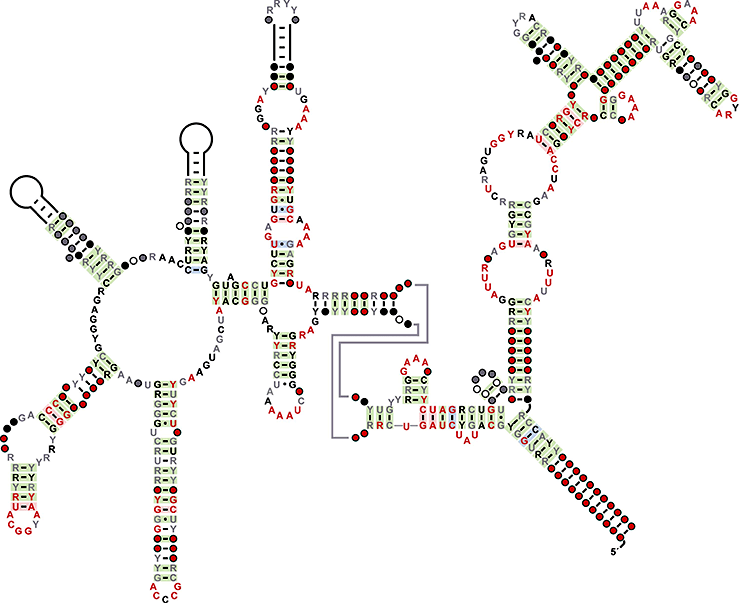
Консенсусная последовательность и вторичная структура OLE РНК, составленная на основе анализа 657 молекул. R — пуриновый нуклеотид, Y — пиримидиновый нуклеотид.
В последние годы длинные некодирующие РНК, или днкРНК (то есть РНК, достигающие более 200 нуклеотидов в длину и, как правило, не содержащие открытых рамок считывания), находятся на пике популярности: каждую неделю выходят свежие публикации, рассказывающие о новых функциях этих, как раньше считалось, бесполезных молекул [1]. Эти «темные лошадки» мира РНК оказались мощнейшими регуляторами экспрессии генов в эукариотических клетках (рис. 1). Заинтересовались ими и медики, так как некоторые длинные некодирующие РНК могут служить биомаркерами различных заболеваний, в том числе онкологических [2]. А есть ли длинные некодирующие РНК у бактерий? Как показывают последние исследования, в прокариотических клетках существует множество классов днкРНК, однако об их биологических функциях практически ничего не известно. В нашем обзоре мы расскажем о тех немногих классах длинных некодирующих РНК бактерий, о которых исследователям удалось что-то узнать.

Рисунок 1. Основные модели функционирования длинных некодирующих РНК. Длинные некодирующие РНК могут связывать определенные белки, не давая им взаимодействовать со своими естественными мишенями и таким образом инактивируя («приманки»). Многие днкРНК выступают в роли структурной основы для сборки белковых комплексов. Другие направляют определенные белки (например, ферменты, модифицирующие гистоны) к ДНК. Наконец, длинные некодирующие РНК могут работать подобно энхансерам, вызывая выпетливание хромосомной ДНК и сближение удаленных друг от друга участков хромосом.
Стоит отметить, что формально под определение длинных некодирующих РНК подпадают некоторые хорошо изученные молекулы, такие как 16S и 23S рРНК, интроны I и II групп, рибозим рибонуклеаза P и транспортно-матричные РНК. В данном обзоре мы их рассматривать не будем, а сконцентрируем внимание на недавно описанных с помощью методов биоинформатики группах длинных некодирующих РНК .
«Биомолекула» не раз писала о днкРНК. Обзорно некодирующие РНК описывает статья «Обо всех РНК на свете, больших и малых» [3]. Материал «Кодирующие некодирующие РНК» [4] рассказывает о пептидах, кодируемых длинными некодирующими РНК. А статья «Загадочное путешествие некодирующей РНК Xist по X-хромосоме» [5] посвящена днкРНК Xist, играющей ключевую роль в инактивации Х-хромосомы у млекопитающих.
Новые классы длинных некодирующих РНК были идентифицированы с помощью биоинформатики, в частности, филогенетическим анализом. Эти РНК имеют длинные, довольно консервативные последовательности и практически не содержат повторов. Более того, они имеют сложную пространственную структуру. Большинство РНК выполняют свои функции, взаимодействуя с другими молекулами нуклеиновых кислот и белков или осуществляя катализ некоторых реакций, и кажется маловероятным, чтобы такие сложные молекулы, как бактериальные длинные некодирующие РНК, были задействованы только в столь простых процессах. Как правило, молекулы РНК со сложной пространственной структурой осуществляют реакции саморазрезания или работают как рибопереключатели, однако все такие молекулы, известные на данный момент, короче 200 нуклеотидов . Вполне возможно, что днкРНК бактерий выполняют функции, ранее не описанные для молекул РНК вообще. Пока же рассмотрим их классы, известные на сегодняшний день.
OLE РНК
OLE (от англ. Ornate, Large, Extremophilic) РНК описаны в 2006 году при анализе 15 бактериальных геномов. На данный момент известно 657 молекул этой группы, причем все они найдены у экстремофильных бактерий или в образцах из окружающей среды. OLE РНК есть у многих представителей бактериального типа фирмикут, и они содержат довольно консервативную последовательность длиной 600 нуклеотидов, которая необходима для формирования сложной вторичной структуры, включающей несколько стеблей (см. заглавный рисунок). Сложная система выпячивания и петель, а также расположение консервативного участка свидетельствуют, что OLE РНК формируют развитую третичную структуру , необходимую для их функционирования.
Так как OLE РНК широко распространены среди экстремофилов, кажется правдоподобным, что они как-то связаны с защитой этих микроорганизмов от агрессивных условий окружающей среды. Анализ транскриптома экстремофильной бактерии Bacillus halodurans показал, что ее ген ole, кодирующий OLE РНК, относится к числу наиболее интенсивно экспрессируемых. Более того, уровень его экспрессии повышается, когда бактерии подвергаются воздействию короткоцепочечных спиртов вроде этанола, которые образуются при росте бактерий в анаэробных условиях.
У многих бактерий ген ole входит в состав крупного оперона, кодирующего белки, участвующие в биосинтезе изопреноидов, репарации ДНК, метаболизме коферментов и регуляции транскрипции. Рядом с геном ole в опероне находится ген белка с неизвестной функцией, известного как OAP (от англ. OLE-associated protein), причем положение обоих генов в опероне относительно друг друга и других генов очень консервативно. Кроме того, OLE РНК локализуется вблизи клеточной мембраны, но только в присутствии OAP. Некоторые исследователи предполагают, что OLE РНК каким-то образом связаны с биосинтезом изопреноидов и, следовательно, влияют на биохимию клеточной мембраны, обеспечивая адаптацию клетки к экстремальным условиям. И как показали эксперименты с нокаутом генов, ни OLE РНК, ни OAP клеткам в нормальных условиях не нужны.
На данный момент OLE РНК составляют наиболее многочисленную группу бактериальных РНК длиной более 500 нуклеотидов, функции которых неизвестны. Более того, OLE РНК — одни из самых консервативных и сложно структурированных некодирующих РНК бактерий.
GOLLD РНК
GOLLD (от англ. Giant, Ornate, Lake- and Lactobacillales-Derived) РНК достигают более 800 нуклеотидов в длину, что делает их третьей по длине группой бактериальных РНК после 16S и 23S рРНК. Как и OLE РНК, GOLLD РНК, по всей видимости, имеют сложную третичную структуру. В большинстве случаев GOLLD РНК кодируются лизогенными бактериофагами, причем их гены нередко соседствуют с генами тРНК. Впрочем, у некоторых бактерий также есть GOLLD РНК, никак не связанные с бактериофагами. Вероятно, GOLLD РНК выполняют какую-то функцию, полезную и для вирусов, и для бактерий или же, напротив, представляют собой эгоистичные генетические элементы.
В пределах последовательности примерно 15% GOLLD РНК имеется тРНК. Смысл такого перекрывания неясен: предполагают, что GOLLD РНК могут каким-то образом участвовать в процессинге тРНК или регулировать их активность. Впрочем, GOLLD РНК и тРНК могут быть и совсем не связаны функционально, а их перекрывание просто позволяет получать сразу две некодирующие РНК из одного предшественника.
Эксперименты с Lactobacillus brevis, у которой ген GOLLD РНК (golld) находится в составе профага, показали, что уровень экспрессии этого гена связан с интенсивностью формирования вирусных частиц. Можно предположить, что GOLLD РНК каким-то образом способствует образованию новых вирионов, однако в клетках с нокаутированным геном golld образование вирусных частиц никак не нарушалось. Таким образом, функции GOLLD РНК остаются загадкой.
HEARO РНК
HEARO (от англ. HNH Endonuclease-Associated RNA and ORF) РНК представляют собой молекулы РНК длиной от 350 нуклеотидов с хорошо развитой пространственной структурой и открытой рамкой считывания. В большинстве случаев она кодирует предполагаемую эндонуклеазу HNH. По всей видимости, в некоторых случаях HEARO РНК транслируются, что было показано экспериментально для одной бактерии.
HEARO РНК обнаружены у бактерий из самых разных групп — фирмикут, протеобактерий, цианобактерий и актинобактерий. А недавно HEARO РНК нашли у археи Methanosarcina mazei. У некоторых бактерий имеется несколько десятков генов hearo. Столь широкое распространение, как полагают ученые, свидетельствует о том, что гены hearo представляют собой мобильные генетические элементы. Однако функции самой HEARO РНК на данный момент не выяснены.
Другие длинные некодирующие РНК бактерий
Для некоторых недавно описанных длинных некодирующих РНК бактерий все-таки удалось с высокой степенью достоверности установить биохимические функции. Примером может служить так называемая T-крупная (T-Large) РНК, которая напоминает самосплайсирующиеся интроны II типа, подвергшиеся круговой пермутации (то есть перевернутые «с ног на голову»). Т-крупная РНК в самом деле способна осуществлять реакции разрыва фосфодиэфирной связи, происходящие при нормальном сплайсинге РНК, и даже использует те же сайты сплайсинга. Однако из-за круговой пермутации сайты сплайсинга расположены в обратном порядке, поэтому экзоны, получающиеся при таком сплайсинге, имеют кольцевую, а не линейную форму, а интроны вырезаются не в виде лассо, а в виде разветвленной структуры, похожей на букву Т. Т-крупные РНК обнаружены у протеобактерий, цианобактерий и представителей типа Deinococcus-Thermus, однако биологическое значение осуществляемого ими необычного сплайсинга неизвестно.
IMES-1 (от англ. Identified in Marine Environmental Sequences) РНК имеют длину около 220 нуклеотидов, многие из которых очень консервативны, а во вторичной структуре образуют один псевдоузел и одно соединение нескольких стеблей. Метатранскриптомные данные, полученные для бактерий из Тихого океана, показали, что в их клетках IMES-1 РНК в пять раз многочисленнее 5S рРНК, однако причины столь высокого уровня экспрессии неясны. Изучение длинных некодирующих РНК этой группы осложняется еще и тем, что более 400 этих молекул обнаружены не в определенных клетках, а в метагеномных образцах.
В метагеноме рубца коров обнаружили еще одну длинную РНК бактериального происхождения — ROOL (от англ. Rumen-Originating, Ornate, Large). Известно 397 таких молекул, относящихся в основном к бактериям порядков Lactobacillales и Clostridiales. Согласно биоинформатическим предсказаниям, ROOL РНК являются одними из наиболее структурно сложных некодирующих РНК, хотя их последовательности не отличаются консервативностью. ROOL РНК немного напоминают GOLLD РНК: они также нередко тесно связаны с генами тРНК, обнаруживаются у бактериофагов и профагов. Можно предположить, что ROOL и GOLLD РНК выполняют схожие функции, однако значительные структурные различия делают эту гипотезу неправдоподобной.
У бактерий типов фирмикут и актинобактерий описано 1347 так называемых raiA РНК, гены которых очень часто находятся вблизи 5′-нетранслируемых областей генов raiA. Белок raiA связывается с рибосомами и замедляет трансляцию в условиях клеточного стресса. Гены raiA РНК иногда обнаруживают выше (upstream) генов, белковые продукты которых локализуются в периплазматическом пространстве и переносят различные вещества. Возможно, в обоих случаях мотив raiA функционирует как рибопереключатель или другой цис-регуляторный элемент, однако потенциальные лиганды, способные с ним взаимодействовать, пока неизвестны.
Сегодня описано около 20 классов бактериальных некодирующих РНК, функции которых еще не установлены. Мы рассмотрели лишь наиболее изученные и широко распространенные из них. Несомненно, дальнейший прогресс в биоинформатике (особенно метагеномике) позволит выявить еще немало групп этих загадочных молекул, а биохимия, структурная биология и биофизика помогут установить их функции в бактериальных клетках.
Источник: biomolecula.ru























